關(guān)于『細(xì)胞自噬』���,你了解多少��?
1.細(xì)胞自噬簡(jiǎn)介
細(xì)胞自噬(autophagy=self-eating)意為自體吞噬���,是真核細(xì)胞在細(xì)胞自噬相關(guān)基因(autophagy related gene,Atg)的調(diào)控下利用溶酶體降解自身細(xì)胞質(zhì)蛋白和受損細(xì)胞器的過(guò)程。細(xì)胞自噬可防止細(xì)胞損傷�����,促進(jìn)細(xì)胞在營(yíng)養(yǎng)缺乏的情況下存活����,并對(duì)細(xì)胞毒性刺激作出反應(yīng)。細(xì)胞自噬包括生理?xiàng)l件下的基礎(chǔ)型細(xì)胞自噬和應(yīng)激條件下的誘導(dǎo)型細(xì)胞自噬。前者是細(xì)胞的自我保護(hù)機(jī)制,有益于細(xì)胞的生長(zhǎng)發(fā)育����,保護(hù)細(xì)胞防止代謝應(yīng)激和氧化損傷,對(duì)維持細(xì)胞內(nèi)穩(wěn)態(tài)以及細(xì)胞產(chǎn)物的合成�、降解和循環(huán)再利用具有重要作用;但細(xì)胞自噬過(guò)度可能導(dǎo)致代謝應(yīng)激�����、降解細(xì)胞成分,甚至引起細(xì)胞死亡等�����。研究表明�����,細(xì)胞自噬能在細(xì)胞穩(wěn)態(tài)�、衰老��、免疫����、腫瘤發(fā)生及神經(jīng)退行性疾病等多種生理病理過(guò)程中發(fā)揮重要作用。
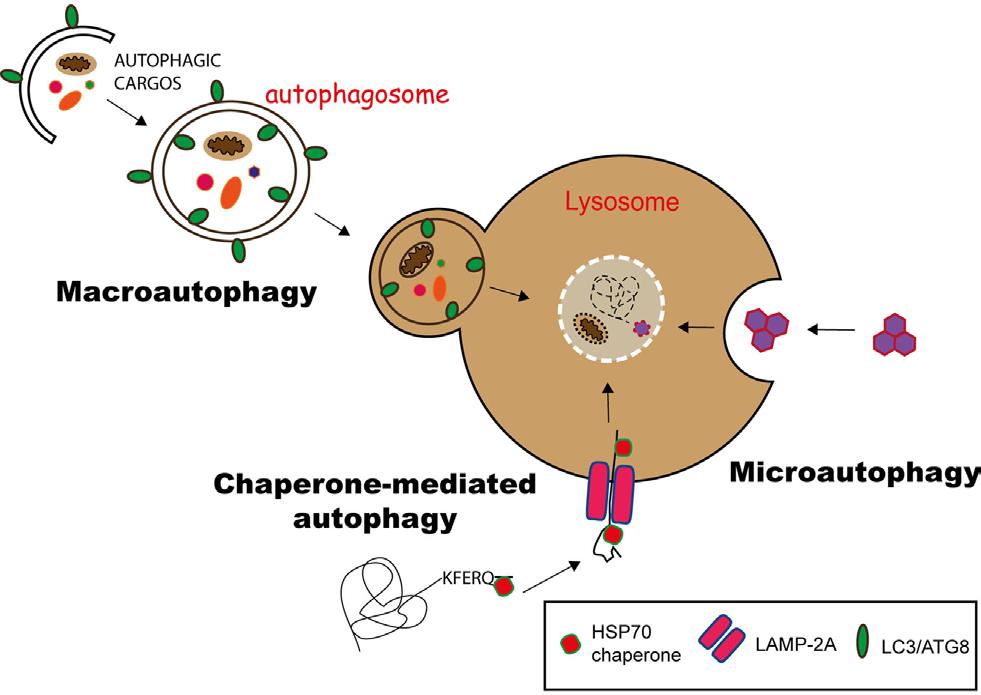
根據(jù)包裹物質(zhì)及運(yùn)送方式的不同可將細(xì)胞自噬分為3種類(lèi)型(圖1):
① 巨細(xì)胞自噬(macroautophagy):通過(guò)形成具有雙層膜結(jié)構(gòu)的細(xì)胞自噬體(autophagosome)包裹胞內(nèi)物質(zhì)��,最終細(xì)胞自噬體與溶酶體融合����。一般情況下所說(shuō)的細(xì)胞自噬是指巨細(xì)胞自噬�。
② 微細(xì)胞自噬(microautophagy):通過(guò)溶酶體或液泡表面的形變直接吞沒(méi)特定的細(xì)胞器��。
③ 分子伴侶介導(dǎo)的細(xì)胞自噬(chaperone-mediated autophagy, CMA):具有KEFRQ樣基序的蛋白在HSP70伴侶的幫助下�,通過(guò)LAMP-2A轉(zhuǎn)運(yùn)體轉(zhuǎn)運(yùn)到溶酶體���。
圖1. 3種細(xì)胞自噬途徑
(Duraes Fernanda V et al. Front Immunol, 2015)
2.細(xì)胞自噬的發(fā)生過(guò)程
細(xì)胞自噬是一種在進(jìn)化上保守的細(xì)胞內(nèi)分解代謝過(guò)程����,在該過(guò)程中�,細(xì)胞質(zhì)大分子���、聚集性蛋白�����、受損細(xì)胞器或病原體被運(yùn)送至溶酶體�,并被溶酶體水解酶降解�����,產(chǎn)生核苷酸����、氨基酸����、脂肪酸、糖和三磷酸腺苷�,最終再循環(huán)到胞漿中�����。諸如饑餓、輻射、缺氧、細(xì)菌入侵、生長(zhǎng)因子匱乏等多種因素均可誘導(dǎo)細(xì)胞自噬發(fā)生。
細(xì)胞自噬的發(fā)生過(guò)程大體分為以下4個(gè)階段(圖2):
①細(xì)胞自噬的起始 在細(xì)胞自噬誘導(dǎo)信號(hào)的調(diào)控下��,ULK1復(fù)合物和多種ATG蛋白被活化�,并定位于前細(xì)胞自噬體處。
②隔離膜和細(xì)胞自噬體的形成 ATG蛋白和脂質(zhì)不斷被募集�����,從而形成杯狀的雙層膜結(jié)構(gòu)(隔離膜�����,phagophore)��;隨著隔離膜的逐漸延伸,將要被降解的胞漿成分完全包裹�,最終形成閉合的細(xì)胞自噬體(autophagosome)�����。
③細(xì)胞自噬體與溶酶體融合 細(xì)胞自噬體形成后將其包裹物通過(guò)胞內(nèi)運(yùn)輸系統(tǒng)運(yùn)輸至溶酶體,并與溶酶體融合���。
④細(xì)胞自噬體的裂解 細(xì)胞自噬體與溶酶體融合后形成細(xì)胞自噬溶酶體(autolysosome)�,最終在溶酶體水解酶的作用下降解其包裹物。
總而言之����,細(xì)胞自噬的本質(zhì)其實(shí)是細(xì)胞內(nèi)的膜重排����。在膜重排的過(guò)程中,會(huì)形成一個(gè)具有雙層膜并裹挾著隨機(jī)或特定底物的封閉囊泡����,即細(xì)胞自噬體,進(jìn)而與溶酶體融合�,形成細(xì)胞自噬溶酶體并降解底物。
圖2. 細(xì)胞自噬的發(fā)生過(guò)程 (Li et al. Molecular Cancer��,2020)
3.細(xì)胞自噬相關(guān)蛋白
細(xì)胞自噬的發(fā)生與營(yíng)養(yǎng)狀態(tài)、能量狀態(tài)��、氧化應(yīng)激�、缺血缺氧等相關(guān)�����,其受多重機(jī)制調(diào)節(jié)��,如ULK1通路���、Beclin1通路����、AMPK通路等����。在細(xì)胞自噬的發(fā)生過(guò)程中,有多種細(xì)胞自噬相關(guān)蛋白可調(diào)節(jié)和控制細(xì)胞自噬形成的不同階段�。
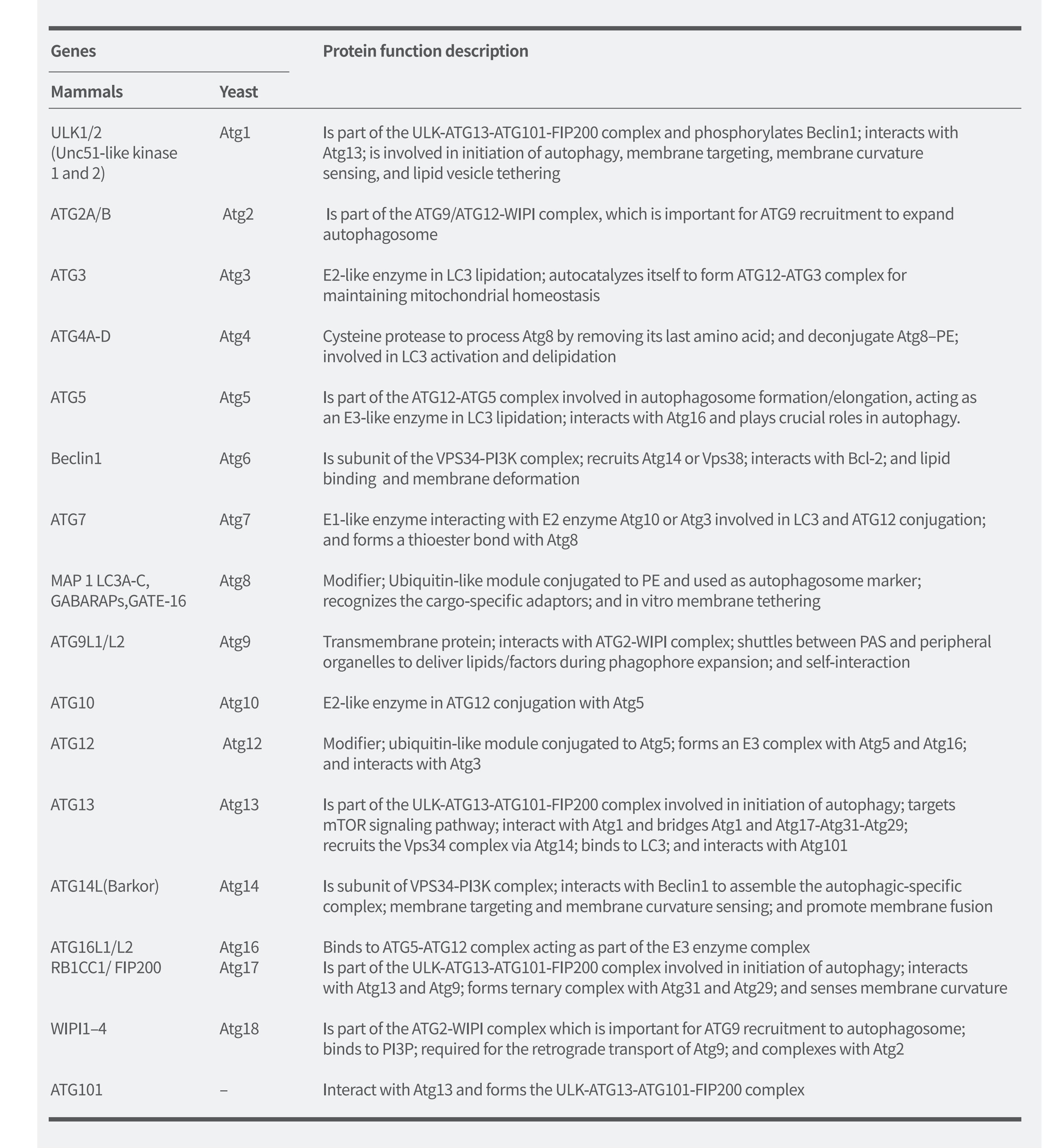
如圖2所示,在細(xì)胞自噬的發(fā)生過(guò)程中,多種細(xì)胞自噬相關(guān)基因調(diào)控細(xì)胞自噬流的不同階段�����。迄今為止���,科學(xué)家已在酵母中鑒定出40余個(gè)編碼ATG蛋白的基因�,并且大多數(shù)在酵母和哺乳動(dòng)物之間高度保守��。在哺乳動(dòng)物細(xì)胞中�����,饑餓誘導(dǎo)的細(xì)胞自噬大約受20種核心ATG基因調(diào)節(jié)�,它們?cè)谝号莞浇徊粩嗄技?����,并組裝形成細(xì)胞自噬前體。這些基因的分類(lèi)和作用(表1)如下:
①ULK1激酶核心復(fù)合物 包括ULK1/2���、ATG13����、RB1CC1/FIP200和 ATG101�;
②PI3K復(fù)合物 包括VPS34、VPS15��、Beclin1和ATG14L;
③ATG9A轉(zhuǎn)運(yùn)系統(tǒng) 包括ATG9A、WIPI1/2和ATG2A;
④ATG12泛素樣結(jié)合系統(tǒng) 包括ATG12、ATG7���、ATG10、ATG5和ATG16L1��;
⑤LC3泛素樣結(jié)合系統(tǒng) 包括LC3A/B/C、ATG7、ATG3和ATG4A/B/C/D����。
表1. 細(xì)胞自噬相關(guān)基因及其在細(xì)胞自噬中的蛋白質(zhì)功能
(Li et al. Molecular Cancer,2020)
4.細(xì)胞自噬的調(diào)控
細(xì)胞基礎(chǔ)水平的細(xì)胞自噬活性很低�����,不適于觀察���,因此,對(duì)細(xì)胞自噬研究多需人工干預(yù)。
雷帕霉素靶蛋白(mTOR)激酶在細(xì)胞自噬反應(yīng)中起著重要的調(diào)節(jié)作用��。mTOR(Akt and MAPK signaling)在被mTOR激酶激活后,細(xì)胞自噬反應(yīng)被抑制;而在mTOR(AMPK and p53 signaling)被抑制后�����,細(xì)胞自噬反應(yīng)機(jī)制啟動(dòng)�����。細(xì)胞自噬相關(guān)基因(autophagy related gene�����,Atg)通過(guò)形成Atg12-Atg5 和 LC3-II (Atg8-II)復(fù)合物來(lái)調(diào)控細(xì)胞自噬體的形成�。
細(xì)胞自噬既能抑制也能促進(jìn)細(xì)胞凋亡�����,兩種反應(yīng)在生物體內(nèi)廣泛存在����。在營(yíng)養(yǎng)缺乏時(shí),細(xì)胞自噬反應(yīng)作為一個(gè)細(xì)胞的促活反應(yīng)機(jī)制存在,但是過(guò)量的細(xì)胞自噬反應(yīng)也會(huì)導(dǎo)致細(xì)胞死亡��,但由細(xì)胞自噬導(dǎo)致的細(xì)胞死亡與細(xì)胞凋亡在形態(tài)學(xué)上有明顯的不同����。幾種促凋亡信號(hào)因子�����,如TNF、TRAIL和 FADD也會(huì)誘導(dǎo)細(xì)胞自噬的反應(yīng)發(fā)生�����。此外���,Bcl-2也是細(xì)胞自噬反應(yīng)重要的調(diào)節(jié)因子,其為凋亡抑制蛋白����,通過(guò)與Beclin-1結(jié)合形成復(fù)合物,來(lái)抑制由Beclin-1誘導(dǎo)的細(xì)胞自噬�����。
已報(bào)道的部分干預(yù)藥物詳見(jiàn)表2:
|
細(xì)胞自噬激動(dòng)劑
|
細(xì)胞自噬拮抗劑
|
|
Rapamycin:雷帕霉素,mTOR抑制劑(最常用)��;
|
Chloroquine:氯奎寧����,溶酶體抑制劑��;
|
|
EBSS:Earle's平衡鹽溶液��,制造饑餓�;
|
Bafilomycin A1:巴佛洛霉素A1��,質(zhì)子泵抑制劑���;
|
|
Lithium Chloride:氯化鋰��,IMPase 抑制劑
|
3-Methyladenine:3-甲基腺嘌呤hVps34抑制劑��;
|
|
Xestospongin B/C:光溜海綿素 B/C���,IP3R阻滯劑等����。
|
NH4Cl:氯化銨,溶酶體抑制劑等�����。
|
表2. 細(xì)胞自噬激動(dòng)劑和拮抗劑
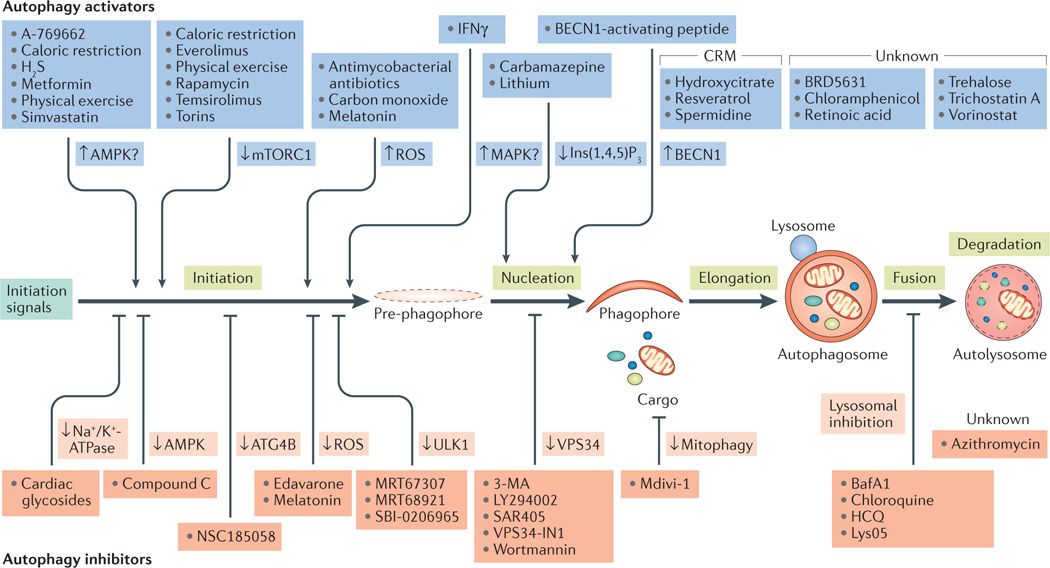
目前常用的細(xì)胞自噬激動(dòng)劑有Rapamycin���,EBSS等��,常用的細(xì)胞自噬抑制劑有Chloroquine,3-MA,NH4Cl和 Bafilomycin A1等。這些細(xì)胞自噬激動(dòng)劑和抑制劑可以分別激活/抑制細(xì)胞自噬發(fā)生的不同階段�,因此研究者可以根據(jù)實(shí)驗(yàn)需要選擇合適的激動(dòng)劑或抑制劑����。更多細(xì)胞自噬通路調(diào)控藥物見(jiàn)圖3。
圖3.細(xì)胞自噬通路調(diào)控藥物
(Lorenzo Galluzi et al. Nat Rev Drug Discov, 2017 )
5.細(xì)胞自噬檢測(cè)方法
在生理?xiàng)l件下��,細(xì)胞自噬活性通常較低。但在饑餓����、缺氧和疾病等刺激下細(xì)胞自噬活性會(huì)顯著上調(diào)�����。另外�����,細(xì)胞自噬抑制也與某些疾病相關(guān)�,如癌癥�、神經(jīng)退行性疾病和傳染病等���。鑒于細(xì)胞自噬與不同的生理和病理生理過(guò)程之間存在緊密聯(lián)系����,故科學(xué)工作者需要選擇理想的細(xì)胞自噬檢測(cè)方法����。細(xì)胞經(jīng)細(xì)胞自噬誘導(dǎo)或抑制后��,常用的觀察和檢測(cè)方法有∶
5.1 透射電鏡法
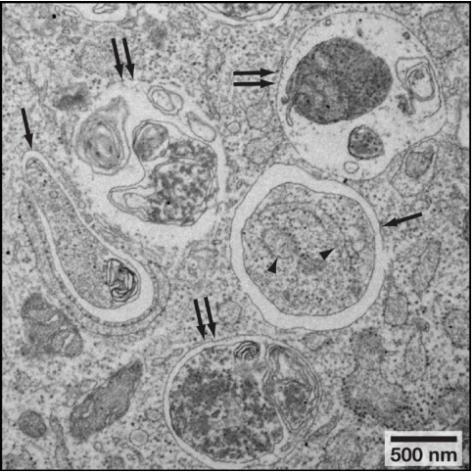
細(xì)胞自噬體屬于亞細(xì)胞結(jié)構(gòu),普通光鏡下看不到�,直接在透射電鏡下觀察細(xì)胞自噬不同階段的形態(tài)變化是一種非常直接的方法��。細(xì)胞自噬各階段的形態(tài)學(xué)特征見(jiàn)表3���;透射電鏡下細(xì)胞自噬小體和細(xì)胞自噬溶酶體的形態(tài)見(jiàn)圖4��。
|
細(xì)胞自噬階段
|
形態(tài)特征
|
|
隔離膜
|
新月?tīng)罨虮瓲?��,雙層或多層膜����,有包繞胞漿成分的趨勢(shì)�。
|
|
細(xì)胞自噬小體
|
雙層或多層膜的液泡狀結(jié)構(gòu),內(nèi)含胞漿成分,如線粒體����、內(nèi)質(zhì)網(wǎng)��、核糖體等。
|
|
細(xì)胞自噬溶酶體
|
單層膜,胞漿成分已降解�。
|
表3. 細(xì)胞自噬各階段特征
圖4. 透射電鏡下細(xì)胞自噬小體(單箭頭)和細(xì)胞自噬溶酶體(雙箭頭)形態(tài)
(Noboru, Mizushima et al. Cell, 2010)
5.2 熒光顯微鏡觀察法
LC3(light chain 3)全稱(chēng)MAP1LC3 (microtubule-associated proteins light chain 3),貫穿整個(gè)細(xì)胞自噬過(guò)程,是目前公認(rèn)的細(xì)胞自噬標(biāo)記物��。哺乳動(dòng)物中的LC3與酵母中的細(xì)胞自噬相關(guān)蛋白Apg8/Aut7/Atg8具有同源性。哺乳動(dòng)物的LC3可分為三種:LC3A����、LC3B和LC3C。其中,LC3B應(yīng)用最為廣泛����。
LC3蛋白合成后在其羧基端被Atg4剪切掉C端5肽�����,暴露甘氨酸殘基�,產(chǎn)生細(xì)胞漿定位的LC3-I�����。在細(xì)胞自噬過(guò)程中�����,LC3-I會(huì)被包括Atg7和Atg3在內(nèi)的泛素樣體系修飾和加工��,與磷脂酰乙醇胺(PE)相偶聯(lián)�����,形成LC3-II并定位于細(xì)胞自噬體內(nèi)外膜上。細(xì)胞自噬體和溶酶體融合后�,外膜上的LC3-II被Atg4切割��,產(chǎn)生LC3-I循環(huán)利用;內(nèi)膜上的LC3-II被溶酶體酶降解���,導(dǎo)致細(xì)胞自噬溶酶體中LC3含量很低(圖5)。因此,科研工作者可以通過(guò)熒光顯微鏡觀察內(nèi)源性LC3或GFP-LC3�����,即可實(shí)現(xiàn)對(duì)細(xì)胞自噬發(fā)生的檢測(cè)�。
圖5. LC3在細(xì)胞自噬發(fā)生中的路徑
(Kiriyama, Y et al. International Electronic Conference on Medicinal Chemistry���,2016)
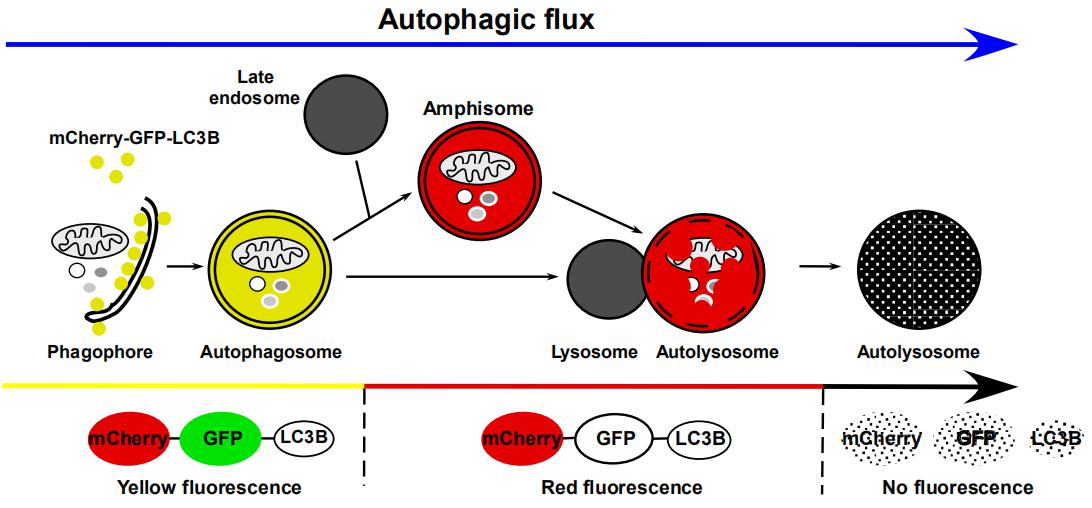
GFP-LC3單熒光和mCherry-GFP-LC3雙熒光指示系統(tǒng)
細(xì)胞自噬形成時(shí)�����,GFP-LC3或mCherry-GFP-LC3融合蛋白轉(zhuǎn)移至細(xì)胞自噬體膜��,在熒光顯微鏡下形成多個(gè)明亮的綠色或黃色熒光斑點(diǎn)。當(dāng)細(xì)胞自噬溶酶體形成后,酸性的溶酶體環(huán)境使GFP熒光淬滅,而mCherry熒光不受影響����,細(xì)胞自噬溶酶體呈現(xiàn)紅色熒光(圖6)��。因此�,科研工作者可以通過(guò)LC3熒光指示系統(tǒng)來(lái)監(jiān)測(cè)細(xì)胞自噬流。
圖6. LC3細(xì)胞自噬雙標(biāo)系統(tǒng)追蹤細(xì)胞自噬流不同階段
(Hansen T E, Johansen T. BMC Biology, 2011)
5.3 Western Blot檢測(cè)LC3和p62蛋白的表達(dá)量
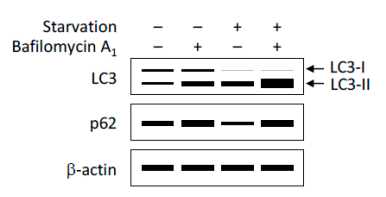
① 利用Western Blot檢測(cè)LC3-II/I比值的變化以評(píng)價(jià)細(xì)胞自噬形成(圖7)�����。細(xì)胞自噬形成時(shí)��,胞漿型LC3-I會(huì)酶解掉一小段多肽�,隨后跟PE結(jié)合轉(zhuǎn)變?yōu)槟ば偷腖C3-II。因此可以通過(guò)LC3-II/I比值的大小估計(jì)細(xì)胞自噬水平的高低。
圖7. WB檢測(cè)LC3����、p62蛋白的表達(dá)結(jié)果示意圖
(Yoshii, S.R. and N. Mizushima. Int J Mol Sci, 2017.)
② 除LC3外��,其他細(xì)胞自噬底物表達(dá)量的變化也可以用于監(jiān)測(cè)細(xì)胞自噬流����。其中,p62是研究廣泛的一個(gè)細(xì)胞自噬底物。p62(也稱(chēng)為SQSTM1蛋白)�,由以下三個(gè)結(jié)構(gòu)域組成:N端Phox和Bem1 (PB1)結(jié)構(gòu)域、鋅指結(jié)構(gòu)域和C端泛素相關(guān)(UBA)結(jié)構(gòu)域�。研究表明,p62蛋白鋅指結(jié)構(gòu)域和UBA結(jié)構(gòu)域之間的連接區(qū)域(LRS區(qū)域)負(fù)責(zé)與細(xì)胞自噬受體蛋白 Atg8/LC3結(jié)合,UBA結(jié)構(gòu)域負(fù)責(zé)招募泛素化蛋白�。在細(xì)胞自噬體形成過(guò)程中����,p62作為鏈接LC3和聚泛素化蛋白之間的橋梁��,被選擇性地包裹進(jìn)細(xì)胞自噬體,之后被細(xì)胞自噬溶酶體中的蛋白水解酶將其降解(圖8)����,所以p62蛋白的表達(dá)量與細(xì)胞自噬活性呈現(xiàn)負(fù)相關(guān)�。因此����,利用Western Blot檢測(cè)p62蛋白的表達(dá)量也可以用來(lái)評(píng)價(jià)細(xì)胞自噬水平(圖7)����。
圖8. p62介導(dǎo)的選擇性細(xì)胞自噬模型
(Ichimura Y et al. J Biol Chem, 2008)
5.4 基于Keima蛋白的細(xì)胞自噬評(píng)價(jià)
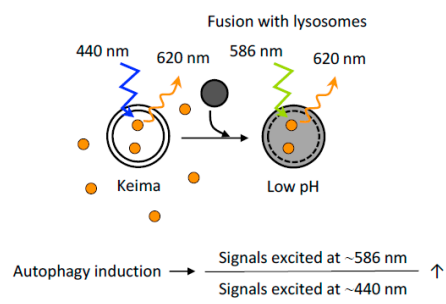
Keima是一種PH敏感型的熒光蛋白���,其雙峰激發(fā)光譜依賴(lài)于周?chē)膒H值�,在中性和酸性環(huán)境中分別在440 nm和586 nm處激活�。Keima在中性和酸性pH中熒光信號(hào)不同的特性�,可以直觀地反映細(xì)胞自噬程度。將細(xì)胞質(zhì)的Keima傳遞到溶酶體可反映非選擇性細(xì)胞自噬���,將Keima融合到特定的蛋白(如融合到線粒體靶向序列—mt-Keima���,作為線粒體細(xì)胞自噬標(biāo)記)可用于反映選擇性細(xì)胞自噬。值得注意是����,基于keima的檢測(cè)不能在固定細(xì)胞中進(jìn)行�����,因?yàn)檫@種檢測(cè)完全依賴(lài)于溶酶體的酸性�。
圖9. 基于Keima蛋白的細(xì)胞自噬評(píng)價(jià)
(Yoshii, S.R. and N. Mizushima. Int J Mol Sci, 2017.)
6.維真細(xì)胞自噬相關(guān)產(chǎn)品
維真生物可提供細(xì)胞自噬研究相關(guān)工具��,包含大量細(xì)胞自噬相關(guān)基因的現(xiàn)貨克隆��,shRNA克隆,CRISPR克隆����,以及LC3細(xì)胞自噬單雙標(biāo)克隆和病毒,多種策略、多種規(guī)格、部分現(xiàn)貨,滿(mǎn)足您多樣化的研究需求�����。
6.1 病毒產(chǎn)品
細(xì)胞自噬單雙標(biāo)病毒
|
產(chǎn)品名稱(chēng)
|
熒光蛋白
|
滴度
|
|
AdV5-CMV-GFP-LC3
|
GFP
|
≥1*10E10pfu/ml
|
|
AdV5-CMV-mCherry-GFP-LC3
|
mCherry�����、GFP
|
≥1*10E10pfu/ml
|
|
AAV-CMV-GFP-LC3
|
GFP
|
≥1*10E13vg/ml
|
|
AAV-CMV-mCherry-GFP-LC3
|
mCherry�����、GFP
|
≥1*10E13vg/ml
|
注:?jiǎn)?dòng)子為CMV
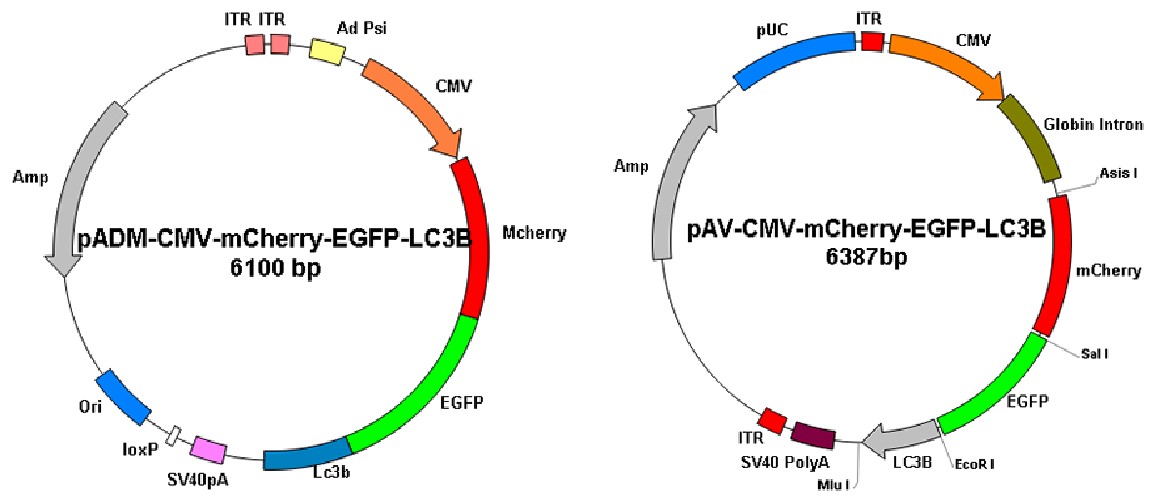
圖10. 維真細(xì)胞自噬雙標(biāo)病毒載體圖譜
6.2 克隆產(chǎn)品
部分細(xì)胞自噬相關(guān)基因
|
產(chǎn)品貨號(hào)
|
基因名稱(chēng)
|
基因編號(hào)
|
基因長(zhǎng)度
|
分類(lèi)
|
|
CH842100
|
ATG13
|
NM_014741
|
1443 bp
|
The ULK1 kinase core complex
|
|
CH870272
|
ATG101
|
NM_021934
|
657bp
|
The ULK1 kinase core complex
|
|
CH826495
|
BECN1/Beclin1
|
NM_003766
|
1353bp
|
The class III PI3K complex I
|
|
CH810399
|
PIK3C3/VPS34
|
NM_002647
|
2664bp
|
The class III PI3K complex I
|
|
CH841329
|
PIK3R4/VPS15
|
NM_014602
|
4077 bp
|
The class III PI3K complex I
|
|
CH838864
|
ATG14/ATG14L
|
NM_014924
|
1479bp
|
The class III PI3K complex I
|
|
CH854934
|
WIPI1
|
NM_017983
|
1341 bp
|
The ATG9A/ATG2-WIPI1/2 trafficking system
|
|
CH882336
|
WIPI2
|
NM_015610
|
1365 bp
|
The ATG9A/ATG2-WIPI1/2 trafficking system
|
|
CH821707
|
ATG12
|
NM_004707
|
423 bp
|
The ATG12-conjugation system
|
|
CH805558
|
ATG10
|
NM_031482
|
663 bp
|
The ATG12-conjugation system
|
|
CH828790
|
ATG5
|
NM_004849
|
828 bp
|
The ATG12-conjugation system
|
|
CH886537
|
ATG16L1
|
NM_030803
|
1824 bp
|
The ATG12-conjugation system
|
|
CH824179
|
MAP1LC3A/LC3A
|
NM_032514
|
366 bp
|
The LC3-conjugation system
|
|
CH837322
|
MAP1LC3B/LC3B
|
NM_022818
|
378 bp
|
The LC3-conjugation system
|
|
CH893675
|
MAP1LC3C/LC3C
|
NM_001004343
|
444 bp
|
The LC3-conjugation system
|
|
CH811236
|
ATG3
|
NM_022488
|
945 bp
|
The LC3-conjugation system
|
|
CH878793
|
ATG4A
|
NM_052936
|
1197 bp
|
The LC3-conjugation system
|
|
CH896350
|
ATG4B
|
NM_013325
|
1182 bp
|
The LC3-conjugation system
|
|
CH804672
|
ATG4C
|
NM_178221/
NM_032852
|
1377 bp
|
The LC3-conjugation system
|
|
CH800065
|
ATG4D
|
NM_032885
|
1425 bp
|
The LC3-conjugation system
|
|
CH871029
|
ATG7
|
NM_006395
|
2112 bp
|
The LC3-conjugation system
|
注:?jiǎn)?dòng)子為CMV���,蛋白標(biāo)簽為C-terminal Flag&6×His
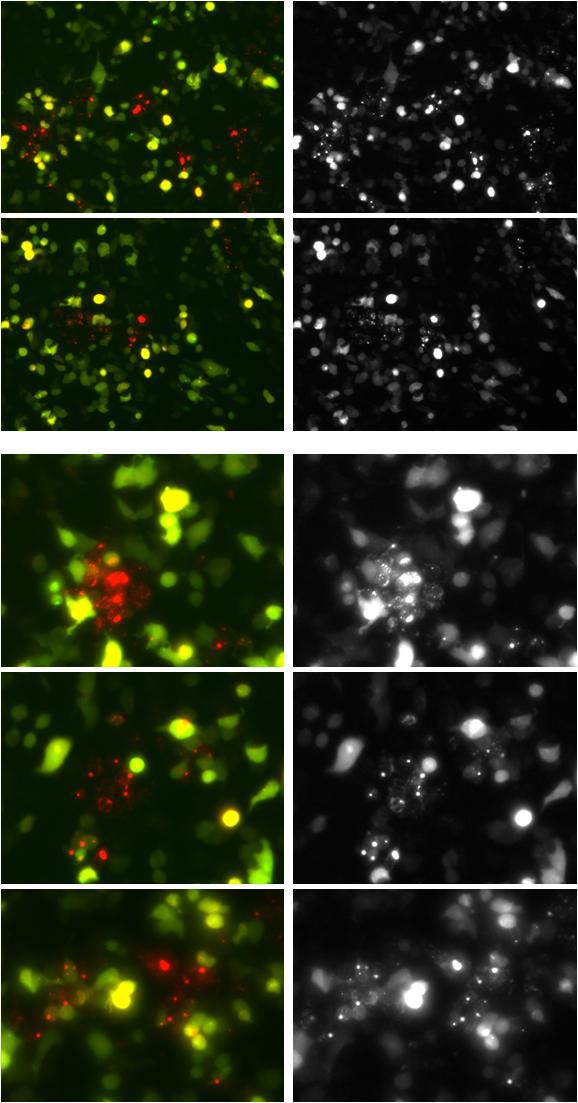
附:維真細(xì)胞自噬雙標(biāo)系統(tǒng)感染誘導(dǎo)細(xì)胞自噬的HEK293細(xì)胞效果圖
7.客戶(hù)代表性文章(部分)
1.Autophagy. (IF=9.77). Peihua Luo, et al. (2020). PLK1 (polo like kinase 1)-dependent autophagy facilitates gefitinib-induced hepatotoxicity by degrading COX6A1(cytochrome coxidase subunit 6A1).[浙江大學(xué) AAV8- shPlk1 & Ad-mCherry-GFP-LC3B 肝功能障礙]
2.Aging (Albany NY). (IF=4.831). Chen Y, et al. (2020). Mitophagy impairment is involved in sevoflurane-induced cognitive dysfunction in aged rats.[浙江大學(xué)醫(yī)學(xué)院 Ad-CMV-mCherry-EGFP-LC3B 術(shù)后認(rèn)知功能障礙]
3.Toxicol Lett. (IF=3.569). Zhang L, et al. (2019). Endoplasmic reticulum stress and autophagy contribute to cadmium-induced cytotoxicity in retinal pigment epithelial cells. [臺(tái)州市第一人民醫(yī)院 AAV-mCherry-eGFP-LC3B 視網(wǎng)膜色素上皮]
4.Molecular Cell. (IF=15.584). Wan, et.al. (2019). Pacer is a mediator of mTORC1 and GSK3-TIP60 signaling in regulation of autophagosome maturation and lipid metabolism.[浙江大AAV9-mCherry-GFP-LC3&AAV9-Pacerwt-HA&Pacer2KR-HA&PacerS157A-HA&PacerS157D-HA 肝細(xì)胞自噬和脂代謝]
5.Cell Physiol Biochem. (IF=5.5). Wang Y, et al. (2018). Inhibition of Histone Deacetylases Prevents Cardiac Remodeling After Myocardial Infarction by Restoring Autophagosome Processing in Cardiac Fibroblasts. [浙江大學(xué)醫(yī)學(xué)院附屬第二醫(yī)院 Ad- mCherry-GFP-LC3 心臟重塑]
6.Biochemical and Biophysical Research Communications. (IF=2.985). Yang, et.al. (2018). Vaspin alleviates myocardial ischaemia/reperfusion injury via activating autophagic ?ux and restoring lysosomal function.[山東大學(xué)齊魯醫(yī)院 AAV-vaspin & Ad-RFP-GFP-LC3 心臟]
7.Mol Cell. (IF=15.584). Wan W, et al. (2018). mTORC1-Regulated and HUWE1-Mediated WIPI2 Degradation Controls Autophagy Flux. [浙江大學(xué)醫(yī)學(xué)院附屬第二醫(yī)院 rAAV-shNC&rAAV-shHUWE1&rAAV-WIPI2-S395&rAAV-WIPI2-S395DAAAV-vaspin 細(xì)胞自噬流]
8.Molecular Cell. (IF=15.584). Su, et al. (2017). VPS34 Acetylation Controls Its Lipid Kinase Activity and the Initiation of Canonical and Non-canonical Autophagy.[浙江大學(xué) AAV9-CMV-VPS34&3KR&3KQ 肝細(xì)胞自噬]