神經(jīng)遞質(zhì)探針
1.研究背景
人類大腦約有860億個神經(jīng)元���,這些神經(jīng)元由數(shù)萬億個突觸連接在一起�。神經(jīng)元的主要功能是接收��、整合和傳遞信息。它們通過電化學(xué)信號進(jìn)行交流�����,并形成神經(jīng)網(wǎng)絡(luò)���,從而控制動物的認(rèn)知和行為����。
神經(jīng)科學(xué)研究的重點�,就是從細(xì)胞和分子水平了解神經(jīng)元如何相互連接和交流,這對于解析動物認(rèn)知和行為背后的神經(jīng)環(huán)路具有重大意義。興奮是神經(jīng)系統(tǒng)信息傳遞的方式����,神經(jīng)纖維受到刺激后�,膜內(nèi)外電位會產(chǎn)生一系列變化而產(chǎn)生興奮�����,興奮產(chǎn)生后先在同一個神經(jīng)元上以動作電位的形式傳導(dǎo),興奮跨過突觸間隙時通過電化學(xué)信號的轉(zhuǎn)變,再傳遞到下一個神經(jīng)元�����。
突觸處有兩種類型的化學(xué)信使,神經(jīng)遞質(zhì)和神經(jīng)調(diào)質(zhì)�����。神經(jīng)遞質(zhì)通過突觸間隙擴(kuò)散��,特異地作用于突觸后細(xì)胞上的受體,從而完成信息傳遞功能��。而神經(jīng)調(diào)質(zhì)則與突觸后細(xì)胞上的受體結(jié)合后��,增強(qiáng)或削弱遞質(zhì)的效應(yīng),從而調(diào)節(jié)突觸信息傳遞��。例如,谷氨酸和γ-氨基丁酸等神經(jīng)遞質(zhì)結(jié)合突觸后細(xì)胞上的受體后,可以迅速地使突觸后神經(jīng)元去極化或超極化,進(jìn)而直接調(diào)控這些神經(jīng)元的活動�。對于神經(jīng)調(diào)質(zhì),它們大多與GPCRs結(jié)合后誘發(fā)突觸前或突觸后電位���,不直接引起突觸后生物學(xué)效應(yīng),但能調(diào)節(jié)遞質(zhì)在突觸前的釋放及突觸后細(xì)胞的興奮性���,調(diào)節(jié)突觸后細(xì)胞對遞質(zhì)的反應(yīng)�。
操控和檢測神經(jīng)元之間的交流是神經(jīng)科學(xué)研究的主要手段,許多遺傳工具已經(jīng)被開發(fā)出來���,這些遺傳工具通過腺相關(guān)病毒(AAV)等病毒載體用于神經(jīng)環(huán)路的操控和成像�����。目前��,光遺傳學(xué)和化學(xué)遺傳學(xué)工具被用來直接操控突觸前神經(jīng)元動作電位的產(chǎn)生�,鈣成像用于檢測突觸后神經(jīng)元內(nèi)的信號傳遞���。然而�,高靈敏度�����、高特異性和高時空分辨率的神經(jīng)遞質(zhì)和神經(jīng)調(diào)質(zhì)檢測技術(shù)還不成熟��。
傳統(tǒng)的神經(jīng)遞質(zhì)檢測技術(shù)����,主要是通過微透析對腦脊液進(jìn)行采樣并生化監(jiān)測��、通過碳纖電極進(jìn)行記錄����,這些檢測手段各自存在諸多弊端,例如特異性差�����、靈敏度低和時空分辨率差,難以精確反映神經(jīng)遞質(zhì)的真實動態(tài)信息等�����。為了解決這一系列的問題���,眾多神經(jīng)科學(xué)家一直在致力于優(yōu)化已有的方法或者開發(fā)新的技術(shù),以求彌補(bǔ)短板,取得突破�。
2.開發(fā)&原理
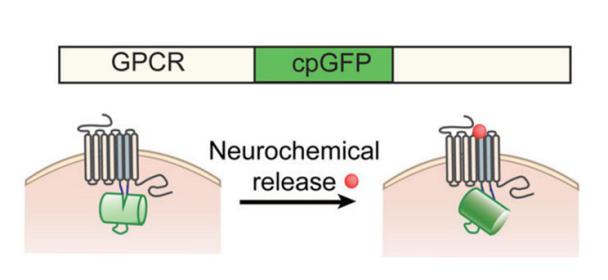
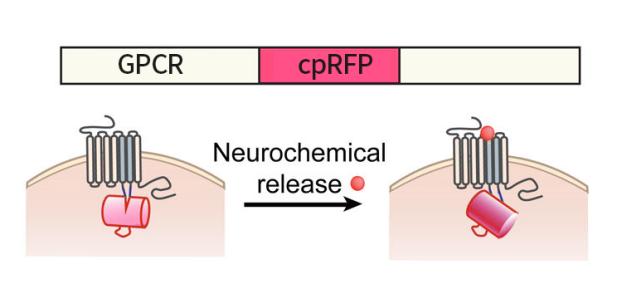
自2018年起,北京大學(xué)李毓龍教授課題組通過偶聯(lián)GPCR和循環(huán)重排熒光蛋白cpFP開發(fā)出了很多檢測神經(jīng)遞質(zhì)的可遺傳編碼的熒光探針�����。神經(jīng)遞質(zhì)與GPCR的結(jié)合會引起后者構(gòu)象的改變��,而這種變化又會引起cpFP發(fā)生構(gòu)象改變����,進(jìn)而影響其發(fā)色團(tuán)周圍的微環(huán)境,最終導(dǎo)致其熒光強(qiáng)度的改變���,這種熒光變化可以通過熒光顯微鏡檢測到(Fig 1a和1b)�。這些可遺傳編碼的熒光探針被命名為GRAB(GPCR Activation-Based)探針����,用于以較高的時空分辨率在體檢測神經(jīng)遞質(zhì)的動態(tài)變化��。
|
|
|
Fig1a:綠色神經(jīng)遞質(zhì)探針(cpEGFP-based)的工作原理
|
Fig1b:紅色神經(jīng)遞質(zhì)探針(cpmApple-based)的工作原理
|
3.探針特征(已發(fā)表)

|
探針名稱
|
所檢測神經(jīng)
化學(xué)物質(zhì)
|
版本
|
顏色
|
骨架
|
親和力
|
信號響應(yīng)幅度
|
動力學(xué)
|
下游
信號偶聯(lián)
|
參考
文獻(xiàn)
|

|
Ach2.0
|
乙酰膽堿
|
第一代
|
綠色
|
人M3受體
|
EC50~1uM
|
ΔF/F0~90%
|
τon~200ms,
τoff~800ms
|
弱偶聯(lián)
|
[8]
|

|
Ach3.0
|
乙酰膽堿
|
第二代
|
綠色
|
人M3受體
|
EC50~2uM
|
ΔF/F0~280%
|
τon~112ms,
τoff~580ms
|
幾乎不偶聯(lián)
|
[3]
|

|
Ach3.0-mut
|
乙酰膽堿
|
第二代對照
|
綠色
|
人M3受體
|
EC50~0uM
(W200A突變)
|
ΔF/F0~1.8%
|
-
|
-
|
[3]
|
|
|
DA1m
|
多巴胺
|
第一代
|
綠色
|
人D2受體
|
EC50~130nM
中等親和力
|
ΔF/F0~90%
|
τon~60ms,
τoff~700ms
|
幾乎不偶聯(lián)
|
[7]
|
|
|
DA1h
|
多巴胺
|
第一代
|
綠色
|
人D2受體
|
EC50~10nM
高親和力
|
ΔF/F0~90%
|
τon~140ms,
τoff~2500ms
|
幾乎不偶聯(lián)
|
[7]
|
|
|
DAmut(1st)
|
多巴胺
|
第一代對照
|
綠色
|
人D2受體
|
EC50~0uM
C118A和S193N突變
|
無效應(yīng)
|
-
|
-
|
[7]
|
|
|
DA2m(DA4.4)
|
多巴胺
|
第二代
|
綠色
|
人D2受體
|
EC50~90nM
中等親和力
|
ΔF/F0~340%
|
τon~40ms,
τoff~1300ms
|
非常小的偶聯(lián)
|
[2]
|
|
|
DA2h(DA4.3)
|
多巴胺
|
第二代
|
綠色
|
人D2受體
|
EC50~7nM
高親和力
|
ΔF/F0~280%
|
τon~50ms,
τoff~7300ms
|
非常小的偶聯(lián)
|
[2]
|
|
|
DAmut(2nd)
|
多巴胺
|
第二代對照
|
綠色
|
人D2受體
|
EC50~0uM
C1183.36A和S1935.42N突變
|
無效應(yīng)
|
-
|
-
|
[2]
|
|
|
rDA1m (rDA2.5m)
|
多巴胺
|
-
|
紅色
|
人D2受體
|
EC50~95nM
中等親和力
|
ΔF/F0~150%
|
τon~80ms,
τoff~770ms
|
非常小的偶聯(lián)
|
[2]
|
|
|
rDA1h (rDA2.5h)
|
多巴胺
|
-
|
紅色
|
人D2受體
|
EC50~4nM
中等親和力
|
ΔF/F0~100%
|
τon~60ms,
τoff~2150ms
|
非常小的偶聯(lián)
|
[2]
|
|
|
rDAmut (rDA2.5mut)
|
多巴胺
|
對照
|
紅色
|
人D2受體
|
EC50~0uM
C1183.36A和S1935.42N突變
|
無效應(yīng)
|
-
|
-
|
[2]
|
|
|
NE1m(NE2.1)
|
去甲腎上腺素
|
-
|
綠色
|
人a2A受體
|
EC50~930nM
中等親和力
|
ΔF/F0~230%
|
τon ~70ms,
τoff~ 750ms
|
不偶聯(lián)
|
[6]
|
|
|
NE1h(NE2.2)
|
去甲腎上腺素
|
-
|
綠色
|
人a2A受體
|
EC50~83nM
高親和力
|
ΔF/F0~130%
|
τon ~30ms,
τoff~ 2000ms
|
不偶聯(lián)
|
[6]
|
|
|
NEmut
|
去甲腎上腺素
|
對照
|
綠色
|
人a2A受體
|
EC50~0uM
S5.46A突變
|
無效應(yīng)
|
-
|
-
|
[6]
|
|
|
Ado1.0
|
腺苷
|
-
|
綠色
|
人A2A受體
|
EC50~60nM
|
ΔF/F0~130%
|
τon~36ms,
τoff~1890ms
|
幾乎不偶聯(lián)
|
[4]
|
|
|
Ado1.0mut
|
腺苷
|
對照
|
綠色
|
人A2A受體
|
EC50~0uM
F168A突變
|
無效應(yīng)
|
-
|
-
|
[4]
|
|
|
5-HT1.0
|
五羥色胺
|
-
|
綠色
|
人5-HT2C
受體
|
EC50~22nM
|
ΔF/F0~250%
|
τon~0.2s,
τoff~3.1s
|
不偶聯(lián)
|
[1]
|
|
|
5-HTmut
|
五羥色胺
|
對照
|
綠色
|
人5-HT2C
受體
|
EC50~0uM
D1343.32Q突變
|
無效應(yīng)
|
-
|
-
|
[1]
|
4.探針資源
在李教授組的授權(quán)下,山東維真生物可以出售這些探針的病毒產(chǎn)品(AAV產(chǎn)品為主)�����,以幫助神經(jīng)科學(xué)工作者在體內(nèi)檢測乙酰膽堿(ACh)���、多巴胺(DA)��、去甲腎上腺素(NE)��、五羥色胺(5-HT)�����、組胺(HA)、腺苷(Ado)����、腺苷三磷酸(ATP)、血管活性腸肽(VIP)�、膽囊收縮素(CCK)�、神經(jīng)肽Y(NPY)���、促腎上腺皮質(zhì)激素釋放因子(CRF)����、精氨酸血管加壓素(AVP)��、催產(chǎn)素(OXT)����、生長激素抑制素(SST)�����、神經(jīng)降壓肽(NTS)���、內(nèi)源性大麻素(eCB)、大麻素(AEA)和褪黑素(MT)等各種遞質(zhì)的動態(tài)變化��,歡迎大家點擊“AAV工具” 中的“神經(jīng)遞質(zhì)探針”進(jìn)行選購!?��。?
5.探針載體獲取
以上所述所有GRAB探針均由北京大學(xué)李毓龍組研發(fā)或聯(lián)合開發(fā)���。部分探針已經(jīng)發(fā)表���,部分探針尚未發(fā)表�。李毓龍組不僅開發(fā)了這些探針�����,還開發(fā)了這些探針的Cre依賴和突變版本�����,包括紅色和綠色的熒光信號,以滿足神經(jīng)科學(xué)工作者多樣化的實驗需求���。此外,為了提升探針的性能�,這些探針還在持續(xù)優(yōu)化中,不斷更新迭代�����。
以上探針均由維真生物協(xié)助提供���。其中已發(fā)表的探針(產(chǎn)品編號中含PUB的病毒為文章中所用病毒),維真生物均有現(xiàn)貨����,當(dāng)天上午下單����,下午/次日即可發(fā)貨;若由于產(chǎn)品脫銷無庫存��,1-2周即可完成入庫并發(fā)貨��;您可以通過公司技術(shù)熱線400-077-2566訂購�。
如若您對未發(fā)表的探針或者探針序列感興趣����,您也可通過上述方式與我們聯(lián)系�,,我們將在第一時間幫您與李教授組溝通���,征得李教授組的同意之后��,我們也可以出售給您。
6.探針病毒使用
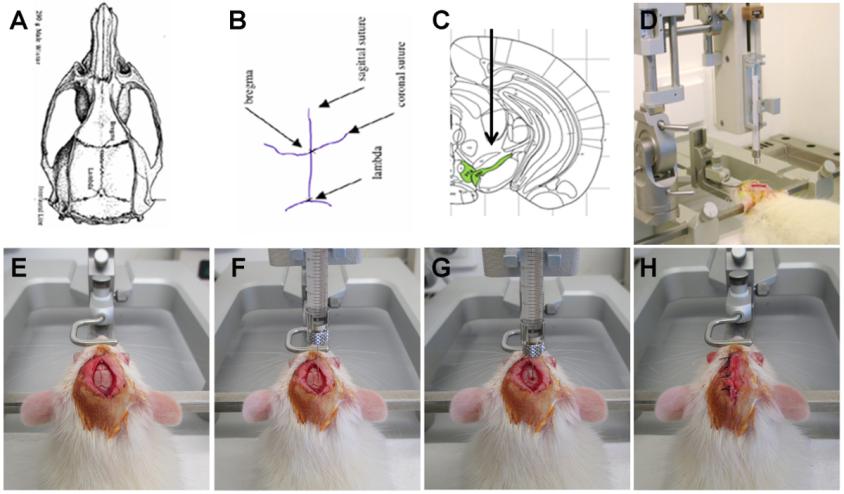
由于血腦屏障的存在�����,普通的注射方式很難實現(xiàn)病毒在腦區(qū)較高且特異性的表達(dá)�,因此研究者通常需要借助腦立體定位儀將探針病毒注射至特定腦區(qū)(詳見Fig 2)�����,以感染全腦和局部腦區(qū)����。
Fig2:腦立體定位注射的示意圖
(Van der Perren, A., et al, V. J. Vis. Exp. (108), e53670, doi:10.3791/53670 (2016)
腦立體定位注射的詳細(xì)步驟如下:
1. 實驗動物稱重�����,進(jìn)行麻醉�����;
2. 待實驗動物完全麻醉后���,用剃毛器將動物頭頂眼睛至耳朵之間的毛發(fā)剃除干凈�;
3. 將麻醉后的實驗動物固定于腦立體定位儀上,具體操作為:實驗動物眼部涂抹青霉素眼藥膏以保護(hù)其雙眼���;將門齒掛在門齒掛鉤上�����,確保頭部保持固定;檢查左右耳桿是否在同一水平上����,將左右耳桿通過外耳道插入實驗動物耳內(nèi)��。實驗動物固定好的標(biāo)準(zhǔn)為:鼻對正中��,頭部不動��,提尾不掉,目測大腦放置水平�����;
4. 手術(shù):用碘伏對實驗動物頭皮進(jìn)行消毒清理��,用手術(shù)刀沿中間位置剪開頭皮�����,用鑷子對表面的結(jié)締組織進(jìn)行清理���,暴露實驗動物的顱骨表面����;然后進(jìn)行調(diào)平,首先找到前囟這個坐標(biāo)�,并將其歸零���,然后向左右調(diào)平�����,使左右兩側(cè)處于同一水平,調(diào)整前后囟�����,使前后囟也在同一水平。
5.病毒注射:通過查詢實驗動物的腦立體定位圖譜確定待注射腦區(qū)的位置(Fig 3a和3b是大鼠和小鼠的腦譜)�����,并確定其坐標(biāo)值���,即ML值(X軸)����、AP值(Y軸)、DV值(Z軸)���;根據(jù)目標(biāo)腦區(qū)設(shè)定好坐標(biāo),移開注射針��,用顱骨鉆開窗(由于顱骨薄,故務(wù)必注意顱骨鉆的力度?。�。僮鲿r避免傷及腦組織;微量注射器吸取病毒液�����,隨后固定在定位儀上,根據(jù)設(shè)好的坐標(biāo)進(jìn)行病毒注射���,注射速度控制在0.1-0.15μl/min�����,注射劑量視具體實驗而定。注射完畢����,留針10min����,以便病毒液充分吸收�,然后慢慢回針。

|
|
|
Fig3a:The Rat Brain
|
Fig3b:The Mice Brain
|
6. 縫合頭皮并消毒, 完成腦定位注射; 將動物從腦立體定位儀取下, 回籠待蘇醒; 動物蘇醒后, 正常喂養(yǎng), 病毒注射后2-4周可檢測轉(zhuǎn)基因的表達(dá)�。
7.常見問題解答
7.1 使用探針AAV病毒注射時���,我該使用多大的病毒量���?
考慮到病毒批次間的差異,我們建議您直接注射未稀釋的病毒原液200-400nl/site����。
7.2 神經(jīng)遞質(zhì)探針在大鼠中是否能像在小鼠中一樣表達(dá)得很好�����?
神經(jīng)遞質(zhì)探針在大鼠中也可以很好地工作�����。根據(jù)李教授組已發(fā)表的論文����,這些探針可以用在多種有機(jī)體中,包括果蠅、斑馬魚���、老鼠和斑馬雀等��。
7.3 你們是否也有廣譜啟動子或其他組織特異性啟動子的探針AAV病毒��?
有些探針在開發(fā)時已構(gòu)建了廣譜啟動子或其他組織特異性啟動子的載體�,如CAG,EFs���,EF1α,CamKIIα,GfaABC1D和TRE等��,具體可以參考探針資源部分的表2��。當(dāng)然��,如果您對其他組織特異性啟動子感興趣��,請參考我們的組織特異性啟動子列表(見下表)來選擇您感興趣的啟動子���。如果您找不到想用的啟動子,可以聯(lián)系我們�����。
|
組織
|
啟動子名稱
|
啟動子
大小
|
啟動子
來源
|
啟動子描述
|
啟動子應(yīng)用
|
|
神經(jīng)
|
hSyn
|
471bp
|
人源
|
突觸蛋白1啟動子
|
神經(jīng)元特異性啟動子
|
|
CamKIIa
|
1.2kb
|
小鼠
|
鈣/鈣調(diào)蛋白依賴性蛋白激酶II α啟動子
|
大腦新皮質(zhì)和海馬興奮性神經(jīng)元特異性啟動子
|
|
c-fos
|
1.7kb
|
小鼠
|
c-fos基因啟動子
|
興奮性神經(jīng)元啟動子
|
|
Mecp2
|
230bp
|
小鼠
|
甲基 CpG 結(jié)合蛋白 2啟動子
|
短的神經(jīng)元特異性啟動子
|
|
NSE
|
1.3kb
|
小鼠
|
烯醇化酶啟動子
|
神經(jīng)元特異性啟動子
|
|
Somatostat(SST)
|
1.2kb
|
人源
|
生長抑制素I基因的啟動子
|
gamma氨基丁酸能抑制性神經(jīng)元SST亞型特異性啟動子
|
|
TH
|
2.5kb
|
大鼠
|
酪氨酸羥化酶基因啟動子
|
多巴胺能神經(jīng)元特異性啟動子
|
|
GFAP
|
2.0kb
|
人源
|
膠質(zhì)纖維酸性蛋白啟動子
|
星形膠質(zhì)細(xì)胞特異性啟動子
|
|
GFAP104
|
845bp
|
人源
|
EF1a和GFAP的嵌合型啟動子
|
星形膠質(zhì)細(xì)胞特異性啟動子
|
|
GfaABC1D(truncated GFAP)
|
681bp
|
人源
|
膠質(zhì)纖維酸性蛋白啟動子
|
星形膠質(zhì)細(xì)胞特異性啟動子
|
|
ALDH1L1
|
1.3kb
|
人源
|
醛脫氫酶1家族成員L1啟動子
|
丘腦中星形膠質(zhì)細(xì)胞特異性啟動子
|
|
MBP
|
1.3kb
|
人源
|
髓磷脂堿性蛋白啟動子
|
少突膠質(zhì)細(xì)胞特異性啟動子
|
|
肝臟
|
ALB
|
2.4kb
|
小鼠
|
白蛋白啟動子
|
肝臟特異性啟動子
|
|
TBG
|
460bp
|
人源
|
甲狀腺素結(jié)合球蛋白啟動子
|
肝臟特異性啟動子
|
|
ApoEHCR-hAAT
|
1.3kb
|
人源
|
載脂蛋白E的肝細(xì)胞控制區(qū)和人α1抗胰蛋白酶啟動子的嵌合啟動子
|
肝臟特異性啟動子
|
|
心臟
|
aMHC
|
0.4kb
|
小鼠
|
肌球蛋白重鏈α啟動子
|
心臟特異性啟動子
|
|
cTNT+intron
|
0.7kb
|
雞
|
心肌肌鈣蛋白T啟動子
|
心肌特異性啟動子
|
|
眼睛
|
Rep65
|
0.7kb
|
小鼠
|
視網(wǎng)膜色素上皮65啟動子
|
視網(wǎng)膜色素上皮細(xì)胞特異性啟動子
|
|
VMD2 promoter
|
0.65bp
|
人源
|
卵黃形成黃斑性營養(yǎng)不良2基因啟動子
|
視網(wǎng)膜色素上皮細(xì)胞特異性啟動子
|
|
胰腺
|
Insulin
|
0.85kb
|
小鼠
|
胰島素基因啟動子
|
胰腺β細(xì)胞特異性啟動子
|
|
PDX1
|
2.7kb
|
小鼠
|
胰十二指腸同源盒1啟動子
|
胰腺β細(xì)胞特異性啟動子
|
|
血管
|
SM22a
|
0.45kb
|
小鼠
|
SM22α啟動子
|
血管平滑肌特異性啟動子
|
|
ICAM2
|
0.15kb
|
人源
|
細(xì)胞間粘附分子2啟動子
|
血管內(nèi)皮特異性啟動子
|
|
CD68
|
0.7kb
|
人源
|
CD68分子啟動子
|
單核巨噬細(xì)胞特異性啟動子
|
|
F4/80
|
1.2kb
|
小鼠
|
F4/80基因promoter
|
巨噬細(xì)胞特異性啟動子
|
|
肌肉
|
MCK
|
1.3kb
|
小鼠
|
肌酸激酶基因啟動子
|
肌肉細(xì)胞特異性啟動子
|
|
3×enhancer McK
|
728bp
|
小鼠
|
修改的肌酸激酶基因啟動子
|
肌肉細(xì)胞特異性啟動子
|
|
腎臟
|
NPHS1
|
1.2kb
|
小鼠
|
Nephrin 1基因啟動子
|
腎臟特異性啟動子
|
7.4 你們探針AAV病毒的最小規(guī)格是多少�����?
我們的探針AAV病毒是按照50μl/支的規(guī)格進(jìn)行分裝的�,因此出售的最小規(guī)格是50μl。
7.5 AAV血清型很多�����,你們有其他血清型的探針AAV病毒嗎,比如逆行AAV血清型���?
對于大多數(shù)神經(jīng)遞質(zhì)探針�,我們通常將其包裝成AAV9血清型����,目前還沒有嘗試逆行AAV血清型。如果您需要逆行血清型�,我們也可以幫您包裝。目前我們可以為您包裝AAV1����,AAV2�����, AAV5���,AAV6�����,AAV7�����,AAV8���,AAVrh10,AAV retro����,AAV ANC80�����,AAV DJ & AAV DJ-8�,AAV PhpB & AAV PhpeB�����,AAV 7m8和AAV shh10等血清型���。
8.參考文獻(xiàn)
8.1 Wan, J., et al. (2021). "A genetically encoded sensor for measuring serotonin dynamics." Nat Neurosci.
8.2 Sun, F., et al. (2020). "Next-generation GRAB sensors for monitoring dopaminergic activity in vivo." Nat Methods 17(11): 1156-1166.
8.3 Jing, M., et al. (2020). "An optimized acetylcholine sensor for monitoring in vivo cholinergic activity." Nat Methods 17(11): 1139-1146.
8.4 Peng, W., et al. (2020). "Regulation of sleep homeostasis mediator adenosine by basal forebrain glutamatergic neurons." Science 369(6508).
8.5 Jing, M., et al. (2019). "G-protein-coupled receptor-based sensors for imaging neurochemicals with high sensitivity and specificity." J Neurochem 151(3): 279-288.
8.6 Feng, J., et al. (2019). "A Genetically Encoded Fluorescent Sensor for Rapid and Specific In Vivo Detection of Norepinephrine." Neuron 102(4): 745-761 e748.
8.7 Sun, F., et al. (2018). "A Genetically Encoded Fluorescent Sensor Enables Rapid and Specific Detection of Dopamine in Flies, Fish, and Mice." Cell 174(2): 481-496 e419.
8.8 Jing, M., et al. (2018). "A genetically encoded fluorescent acetylcholine indicator for in vitro and in vivo studies." Nat Biotechnol 36(8): 726-737.